lunes, 11 de agosto de 2014
domingo, 10 de agosto de 2014
Actividad 1 "Reflexión del artículo"
Los estudiantes en línea son mucho más complejos que los
estudiantes regulares, por lo general son personas mayores que tienen
necesidades diferentes orientadas a mejorar en sus trabajos, la mayoría de
ellos son adultos mayores que trabajan y que tienen que migrar de ambientes
personales a ambientes virtuales y que muchas veces no saben ocupar una
computadora.
Por otro lado la evaluación de un estudiante siempre
pretende medir los conocimientos adquiridos y para ello utilizamos diferentes
técnicas. Dentro del aprendizaje en línea esta labor tiene ventajas y
desventajas (mencionadas en el texto).
La principal ventaja es que el estudiante es la persona que
se hace cargo de su propia evaluación es decir, él entra a las plataformas
virtuales cuando lo necesita, aprende y repasa lo que quiere y el profesor solamente
le proporciona las herramientas necesarias para su guía, al final el alumno es
quien determina si el curso le sirve de algo si resuelve sus necesidades y
cumple sus expectativas.
Esto implica un nivel superior de aprendizaje por parte del
alumno, ya que su nivel de aprendizaje no se limita a la simple retención de
memoria o a la repetición mecánica; ya requiere un nivel de análisis y de
discusión mucho mayor, que no siempre está acostumbrado a utilizar.
Es por ello que el profesor debe mantener una guía clara,
con un principio y un final, donde la evaluación muestra cuales son los parámetros
que se deben cumplir y al mismo tiempo los objetivos que se necesitan para que
el alumno avance.
Tanto el profesor como el alumno deben ser claros, tanto en
el momento de escribir como en el momento de organizar sus ideas y por ello los
métodos de evaluación suelen ser diversos, eso es una enorme ventaja para el
profesor ya que puede organizar diferentes técnicas de evaluación y el estudiante
recibe una retroalimentación inmediata.

Para finalizar la reflexión sería que estos medios y
espacios están generando una revolución en el campo de la educación, cientos de
personas se están avocando al desarrollo de programas de estudio, software,
técnicas, dinámicas y un sinfín de recursos y aplicaciones para que cada vez
más gente cuente con un espacio donde poder estudiar, comprender y manejar
información; desde luego esta visión tecnológica no es 100% fiable, no es
infalible y desde luego que carece de ciertos componentes que están presentes
en las clases presenciales. Pero ante la constante demanda de cursos,
profesores y escuelas de calidad, el aumento de la población estudiantil y la
competencia global esto se convierte en una herramienta más que ayuda a muchas
personas.
miércoles, 6 de agosto de 2014
Actividad 3 "Investigación"

- Web 2.0,
- Moodle
- Blackboard
- Wiki
- Hootcourse
- RSS
- Podcast
- Vodcast
- Glogster
- WebQuest,
- Sites (Google)
Les anexo un documento que me llegó a través de un curso de Laureate donde se presentan algunos de los conceptos presentados arriba pero también otros que podrían ser importantes.
Les anexo un manual de blackboard; yo lo he usado y es increíble se los recomiendo
Manual de Blackboard
Les anexo un manual de blackboard; yo lo he usado y es increíble se los recomiendo
Manual de Blackboard
Herramientas para el
estudiante del siglo 21
Hay una gran variedad de herramientas
disponibles para que los instructores utilicen en sus clases cara a cara y
cursos en línea. Estas herramientas permiten que los estudiantes participen en
su propio aprendizaje y colaboren con sus compañeros. Estas herramientas varían
en diversidad, capacidad, accesibilidad y facilidad de uso. Mientras que las
nuevas herramientas están apareciendo constantemente, este recurso incluye una
lista básica de las tres categorías de herramientas: Herramientas para la
creación, el establecimiento de redes y el intercambio de información.
Herramientas para la
creación
Apps – los Apps permiten
que los estudiantes e instructores desarrollen contenido para una variedad de
dispositivos móviles. Vienen en una gama de variedad, costo y dificultad.
También, los Apps se pueden utilizar para formar contactos y compartir información.
Animoto – los estudiantes y
los maestros pueden fácilmente crear películas y videos que incorporan diversos
medios de comunicación para presentar sus ideas a su comunidad de aprendizaje.
Glogster – una herramienta Web
2.0 que permite crear carteles virtuales que combinan texto, audio, video,
imágenes e hipervínculos, y permite compartirlas con los demás por medios
electrónicos.
MindMeister –software en línea de
mapas conceptuales que les permite a los usuarios crear diagramas al conectar
ideas principales con un tema central.
Prezi – una herramienta de
presentación no lineal que permite incrustar otras formas de multimedia en la
presentación.
SlideRocket – una herramienta en
línea para la creación de presentaciones que les permite a los usuarios crear
presentaciones visualmente atractivas y ricas en multimedia.
VoiceThread – abarca todas las
formas de medios de comunicación en una presentación que invita las respuestas
(en todas las formas de medios de comunicación) de los espectadores.
Sitios para la
creación de sitios web – sitios web que le ayudan a una persona crear un sitio
web mediante un proceso de paso a paso. Adicionalmente, la mayoría de estos
sitios proveen alojamiento para los sitios web por un precio.
Software para la
creación de sitios web – tipos de software que permiten el desarrollo de los
sitios web simples a los complejos. Vienen en una gama de variedad, costo y
dificultad. A menudo, estos se compran a través del Internet o en su tienda
local de tecnología. En general, este tipo de software sólo es compatible con
el desarrollo de un sitio web y le deja al usuario la responsabilidad de
localizar un lugar en el Internet para alojar el sitio web.
Sitios para
tutoriales de sitios web - sitios web que proveen instrucciones para crear un
sitio web. No proveen la creación ni el alojamiento, sino que son sitios educacionales
utilizados para compartir información.
Herramientas para el
establecimiento de redes
IM (Mensajería
Instantánea) –
con mayor frecuencia se refiere al intercambio de mensajes de texto en tiempo
real entre dos o más personas, sin embargo los archivos y los videos también se
pueden intercambiar.
Facebook – se podría decir que
es la red social más conocida en el mundo. Fue diseñada para servir el mismo
propósito que la versión en papel del “Facebook” de los universitarios, una vez
dada a los estudiantes en su primer año para ayudarles a conectar nombres y
rostros. Esta plataforma gratuita permite que los individuos intercambien
palabras, imágenes y enlaces con otros que tengan una cuenta de Facebook.
HootCourse – una red gratuita de
salón en línea que utiliza la aplicación social actualmente preferida por los
estudiantes y los instructores, es decir, Twitter con el que se conectan a la
comunidad del curso.
Twitter – sistema de
mensajería instantánea que permite la comunicación a través del intercambio de
140 caracteres o menos. Originalmente diseñado como una red social, también es
utilizada por los negocios para la publicidad y por las instituciones
educacionales para publicar anuncios para la comunidad de aprendizaje.
Skype – un servicio de
video y teléfono basado en la Web. Skype permite llamadas de computadora a
computadora y llamadas telefónicas a través del Internet.
Herramientas para el
intercambio de información
Adobe Connect – software que
permite las conferencias basadas en la web (webinars) y el aprendizaje
electrónico (eLearning).
Blog – abreviatura de
"weblog", es un diario en línea publicada en orden cronológico
inverso. Debido a que sólo se requiere un mínimo conocimiento de computadoras
para publicar una página de blog, el acto de "blogging" ha creado
una nueva avenida para la escritura. Además de pensamientos y reacciones
personales, profesionales suelen promover el “blogging” como un medio de
intercambio de información.
Google Docs – un grupo de
aplicaciones basadas en la Web, tales como procesamiento de textos, hoja de
cálculo, presentaciones, creación de formularios, y el almacenamiento en línea
que permiten a los usuarios cargar documentos en la mayoría de las formas y
descargar en los demás.
Microsoft SkyDrive – un sitio de
almacenamiento en línea que permite a los usuarios subir fotos y documentos que
se puede acceder desde otras computadoras y teléfonos.
RSS (Really Simple
Syndication) Feeds –
un sistema para la recopilación de blog y actualizaciones de sitios de noticias
e información.
TeacherTube – sitio web para los
educadores para compartir videos. Le permite a cualquiera subir videos cortos
para la observación privada o pública. Es un sitio web acompañante de YouTube.
Vodcast/Podcast – un clip de video
para ser visto a través de un dispositivo portátil. Podcast: un archivo mp3 (o
formato similar) que se puede oír a través de un reproductor de música digital.
VoiceThread – abarca todas las
formas de medios de comunicación en una presentación que invita las respuestas
(en todas las formas de medios de comunicación) de los espectadores.
Workflowy – una herramienta de
colaboración en línea que les permite a los usuarios organizar listas,
planificar proyectos, tomar notas, escribir trabajos de investigación y más.
Webinars – un seminario basado
en la Web presentada a través del Internet. Webinars pueden ser interactivos o
transmisiones unidireccionales.
Wiki – derivado de la
expresión hawaiana "wiki wiki", que significa
"súperrápido". Un wiki es un servicio web que permite a los usuarios
añadir y actualizar el contenido del sitio web. Esto crea una obra de
colaboración que está en constante evolución.
Actividad 4 "Membrana Plasmática"
MEMBRANA PLASMÁTICA.

La membrana plasmática fue definida en 1967 por Palade como un complejo molecular que delimita un territorio celular determinado. Durante mucho tiempo sólo se pudo intuir su existencia ya que es una estructura tan delgada que escapaba a la observación mediante el microscopio óptico. Hoy sabemos que la membrana plasmática es una envoltura continua que rodea la célula estando una de sus caras en contacto con el medio extracelular y la otra con el hialoplasma. La observación al microscopio electrónico revela una estructura de unos 7 nm de grosor en la que se aprecian dos bandas oscuras separadas por una banda más clara. La Figura 11.3 contiene una fotografía al microscopio electrónico en la que se aprecia el aspecto trilaminar de las membranas plasmáticas de dos neuronas vecinas y la hendidura sináptica que las separa
La casi totalidad de la masa de la membrana plasmática está constituida por proteínas y lípidos anfipáticos; contiene además pequeñas cantidades de glúcidos en forma de oligosacáridos unidos covalentemente a las proteínas o a los lípidos. La membrana plasmática del eritrocito humano (una de las mejor estudiadas) está formada por aproximadamente un 60% de proteínas y un 40% de lípidos. Estas proporciones pueden variar de acuerdo con el tipo de célula, pero en la mayor parte de los casos la masa total de proteínas supera a la de lípidos. Las membranas que constituyen el sistema membranario interno característico de la célula eucariota presentan una composición y estructura muy similares a las de la membrana plasmática.
Los lípidos que aparecen formando parte de las membranas biológicas son fosfoglicéridos, esfingolípidos ycolesterol (u otros esteroles afines). Todos ellos tienen en común su carácter marcadamente anfipático que los hace idóneos para este cometido. Las proporciones relativas de los distintos tipos de lípidos varían según el tipo de célula; las membranas presentes en cada reino, especie, tejido y tipo celular poseen una composición lipídica característica, e incluso, dentro de un mismo tipo celular esta composición puede variar entre los diferentes orgánulos. Está claro que las células poseen mecanismos para regular la composición lipídica de sus membranas y que las diferencias existentes en este aspecto deben tener algún significado funcional, aunque en la mayor parte de los casos está por descubrir cual es.
La composición proteica de membranas biológicas de orígenes diferentes varía aún más ampliamente que su composición lipídica, lo que refleja que estas proteínas deben estar especializadas en determinadas funciones que son diferentes según el tipo de célula. Como regla general, una membrana biológica posee varios centenares de proteínas diferentes, la mayoría de las cuales están especializadas en el transporte de solutos específicos a su través.
Todas las membranas biológicas comparten ciertas propiedades fundamentales. Son poco permeables a los solutos cargados o polares pero permeables a las sustancias apolares; tienen un grosor de entre 5 y 8 nm y apariencia trilaminar (dos bandas oscuras separadas por una banda clara) cuando se las examina en sección transversal con el microscopio electrónico. La combinación de las pruebas obtenidas mediante microscopía electrónica, el análisis de la composición química, y los estudios físicos realizados acerca de la permeabilidad de las membranas biológicas y de la movilidad de las moléculas individuales de lípidos y proteínas que las constituyen, fructificaron a comienzos de los años 70 en el modelo del mosaico fluido, que explica la estructura de dichas membranas. Según este modelo, la estructura básica de la membrana es una bicapa lipídica formada por lípidos anfipáticos en la que las porciones apolares de dichos lípidos se encuentran encaradas unas con otras en el centro de la bicapa y sus grupos de cabeza polares encarados hacia el exterior a ambos lados de la misma. Las proteínas, que son de tipo globular, se encuentran incrustadas a intervalos irregulares en la bicapa manteniéndose unidas a ella mediante interacciones hidrofóbicas entre sus zonas apolares y las zonas apolares de los lípidos. La estructura es fluida, es decir, las moléculas individuales de lípidos y proteínas, debido a que se mantienen unidas por interacciones no covalentes, tienen libertad para moverse lateralmente en el plano de la membrana. La cara externa de la membrana plasmática, la que da al medio extracelular, presenta cadenas oligosacarídicas unidas covalentemente a lípidos o a proteínas; otras membranas celulares no presentan estos componentes glucídicos. En la Figura 11.4 se representa esquemáticamente el modelo del mosaico fluido para la estructura de la membrana plasmática.

El modelo del mosaico fluido explica la apariencia trilaminar de las membranas cuando se observan al microscopio electrónico: las dos bandas oscuras externas corresponderían a los grupos de cabeza polares (que contienen átomos más pesados y que por lo tanto retienen más los electrones), mientras que la banda clara correspondería a las colas no polares de los lípidos de membrana (formadas por átomos más ligeros y por lo tanto más transparentes a los electrones). Por otra parte, un grosor de entre 5 y 8 nm es el que cabría esperar de una bicapa de lípidos anfipáticos con proteínas incrustadas en ella y sobresaliendo a ambos lados. Los experimentos llevados a cabo con diferentes tipos de lípidos anfipáticos indican que éstos en medio acuoso tienden espontáneamente a formar estructuras como las micelas, bicapas y liposomas, ya estudiadas anteriormente, lo que también apoya la idea de una bicapa lipídica como elemento básico estructural en las membranas celulares.
El carácter fluido de las membranas se estableció mediante el marcaje de moléculas individuales de lípidos y su posterior seguimiento en el seno de la bicapa. Así pudo comprobarse que los movimientos de los lípidos dentro de cada monocapa son muy frecuentes y rápidos, mientras que son mucho más raros los cambios de una a otra monocapa (difusión "flip-flop").
El grado de fluidez de las membranas depende de la temperatura y de la composición en ácidos grasos de sus lípidos constituyentes. A temperaturas muy bajas los lípidos de membrana tienden a adoptar un ordenamiento casi cristalino (paracristalino). Por encima de una temperatura que es característica de cada membrana las moléculas de los lípidos comienzan a moverse y la membrana pasa al estado fluido. La temperatura de transición del estado paracristalino al estado fluido depende de la composición lipídica; los ácidos grasos saturados favorecen un mayor empaquetamiento de los lípidos de membrana y por lo tanto la temperatura de transición será mayor cuanto mayor sea la proporción de éstos ácidos grasos en la membrana. Por el contrario, los cambios de orientación existentes en las cadenas de los ácidos grasos insaturados dificultan el empaquetamiento de los lípidos de membrana en un ordenamiento paracristalino, por lo tanto la temperatura de transición será menor cuanto más abundante sea este tipo de ácidos grasos. Esta es la razón por la que en los organismos homeotermos, que mantienen una temperatura corporal constante y elevada, abundan los lípidos de membrana ricos en ácidos grasos saturados, mientras que en los organismos poiquilotermos, incapaces de regular su temperatura corporal, son más abundantes los lípidos de membrana ricos en ácidos grasos insaturados. Se ha comprobado que distintos tipos de células mantenidos en cultivo son capaces de alterar la composición en ácidos grasos de sus lípidos de membrana respondiendo a las variaciones de la temperatura ambiente, todo ello con el objeto de mantener la fluidez de sus membranas celulares. Por otra parte, el colesterol y esteroles afines, debido a la rigidez del sistema de anillos condensados de su molécula, tienden a impedir, actuando a modo de cuña, la agregación de los demás lípidos de membrana en ordenamientos paracristalinos, con lo que también colaboran en el mantenimiento del estado fluido (Figura 11.4).

El carácter fluido de las membranas es de una gran importancia biológica. Por una parte este carácter fluido permite que la membrana, y con ella la célula, sufra deformaciones que son la base de muchos movimientos celulares. Por otra, posibilita que tanto la membrana plasmática, como las de diferentes orgánulos puedan sufrir fenómenos de fusión y escisión del tipo de los que se representan en la Figura 11.5; estos fenómenos, en los que porciones de membrana se desprenden en forma de vesículas para luego fundirse con otras membranas, constituyen la base la base de una compleja relación dinámica entre los distintos compartimentos celulares que se conoce con el nombre de flujo de membrana y será estudiada con más detalle en otro capítulo.
En cuanto a las proteínas que forman parte de la membrana se distinguen dos tipos según su mayor o menor grado de asociación con la bicapa lipídica (ver Figura 11.6):
1) Proteínas integrales (o intrínsecas).- Se encuentran íntimamente asociadas a la bicapa por lo que resulten difíciles de extraer de la misma. Una parte sustancial de su molécula se encuentra sumergida en la bicapa lipídica estableciéndose interacciones hidrofóbicas entre los grupos R de los restos de aminoácidos no polares y las colas hidrocarbonadas de los lípidos. Se pueden extraer de la bicapa utilizando detergentes, que interfieren con dichas interacciones hidrofóbicas; cuando se realiza la extracción tienden a precipitar una vez eliminado el detergente, es decir, son poco solubles en agua. Algunas proteínas integrales atraviesan la membrana de lado a lado (proteínas transmembrana) y otras lo hacen sólo en parte.
En los últimos años se ha podido determinar la conformación tridimensional de unas cuantas proteínas integrales; todas ellas presentan dos dominios bien diferenciados: en uno de ellos, el que se encuentra sumergido en la bicapa, los restos de aminoácidos hidrofóbicos se sitúan exteriormente, de manera que puedan interactuar con las colas hidrocarbonadas de los lípidos; en el otro, formado por las zonas de la molécula que asoman a uno y otro lado de la bicapa, son los restos de aminoácidos polares los que se encuentran situados en superficie, de manera que quedan expuestos al agua. Este tipo de conformación tridimensional contrasta con el mucho más común que presentan las proteínas globulares solubles, las cuales tienen todos sus restos polares en superficie y los no polares sepultados en el núcleo proteico.ico.
2) Proteínas periféricas (o extrínsecas).- Su grado de asociación con la bicapa es mucho más débil. Se encuentran unidas a las cabezas polares de los lípidos o bien a proteínas integrales mediante interacciones débiles. Pueden liberarse de la membrana con tratamientos relativamente suaves (cambios en el pH o fuerza iónica) que rompen estas interacciones; una vez liberadas son completamente hidrosolubles. Algunas proteínas periféricas están ancladas covalentemente a lípidos de la bicapa y pueden ser liberadas por acción de enzimas específicos. Las proteínas periféricas presentan conformaciones tridimensionales que en general no difieren de las de otras proteínas globulares.
La composición y estructura supramolecular de las membranas celulares, que acabamos de estudiar, hacen que éstas, lejos de constituir meras barreras pasivas, participen de una manera activa en la regulación del tráfico de sustancias entre la célula y su entorno y entre los distintos compartimentos celulares.
Tomado de CURSO DE BIOLOGÍA Alejandro Porto Andión http://www.bionova.org.es/biocast/p2temas.htm todos los derechos reservados, se copia con fines de educación y sin fines de lucro
La membrana plasmática fue definida en 1967 por Palade como un complejo molecular que delimita un territorio celular determinado. Durante mucho tiempo sólo se pudo intuir su existencia ya que es una estructura tan delgada que escapaba a la observación mediante el microscopio óptico. Hoy sabemos que la membrana plasmática es una envoltura continua que rodea la célula estando una de sus caras en contacto con el medio extracelular y la otra con el hialoplasma. La observación al microscopio electrónico revela una estructura de unos 7 nm de grosor en la que se aprecian dos bandas oscuras separadas por una banda más clara. La Figura 11.3 contiene una fotografía al microscopio electrónico en la que se aprecia el aspecto trilaminar de las membranas plasmáticas de dos neuronas vecinas y la hendidura sináptica que las separa
La casi totalidad de la masa de la membrana plasmática está constituida por proteínas y lípidos anfipáticos; contiene además pequeñas cantidades de glúcidos en forma de oligosacáridos unidos covalentemente a las proteínas o a los lípidos. La membrana plasmática del eritrocito humano (una de las mejor estudiadas) está formada por aproximadamente un 60% de proteínas y un 40% de lípidos. Estas proporciones pueden variar de acuerdo con el tipo de célula, pero en la mayor parte de los casos la masa total de proteínas supera a la de lípidos. Las membranas que constituyen el sistema membranario interno característico de la célula eucariota presentan una composición y estructura muy similares a las de la membrana plasmática.
Los lípidos que aparecen formando parte de las membranas biológicas son fosfoglicéridos, esfingolípidos ycolesterol (u otros esteroles afines). Todos ellos tienen en común su carácter marcadamente anfipático que los hace idóneos para este cometido. Las proporciones relativas de los distintos tipos de lípidos varían según el tipo de célula; las membranas presentes en cada reino, especie, tejido y tipo celular poseen una composición lipídica característica, e incluso, dentro de un mismo tipo celular esta composición puede variar entre los diferentes orgánulos. Está claro que las células poseen mecanismos para regular la composición lipídica de sus membranas y que las diferencias existentes en este aspecto deben tener algún significado funcional, aunque en la mayor parte de los casos está por descubrir cual es.
La composición proteica de membranas biológicas de orígenes diferentes varía aún más ampliamente que su composición lipídica, lo que refleja que estas proteínas deben estar especializadas en determinadas funciones que son diferentes según el tipo de célula. Como regla general, una membrana biológica posee varios centenares de proteínas diferentes, la mayoría de las cuales están especializadas en el transporte de solutos específicos a su través.
Todas las membranas biológicas comparten ciertas propiedades fundamentales. Son poco permeables a los solutos cargados o polares pero permeables a las sustancias apolares; tienen un grosor de entre 5 y 8 nm y apariencia trilaminar (dos bandas oscuras separadas por una banda clara) cuando se las examina en sección transversal con el microscopio electrónico. La combinación de las pruebas obtenidas mediante microscopía electrónica, el análisis de la composición química, y los estudios físicos realizados acerca de la permeabilidad de las membranas biológicas y de la movilidad de las moléculas individuales de lípidos y proteínas que las constituyen, fructificaron a comienzos de los años 70 en el modelo del mosaico fluido, que explica la estructura de dichas membranas. Según este modelo, la estructura básica de la membrana es una bicapa lipídica formada por lípidos anfipáticos en la que las porciones apolares de dichos lípidos se encuentran encaradas unas con otras en el centro de la bicapa y sus grupos de cabeza polares encarados hacia el exterior a ambos lados de la misma. Las proteínas, que son de tipo globular, se encuentran incrustadas a intervalos irregulares en la bicapa manteniéndose unidas a ella mediante interacciones hidrofóbicas entre sus zonas apolares y las zonas apolares de los lípidos. La estructura es fluida, es decir, las moléculas individuales de lípidos y proteínas, debido a que se mantienen unidas por interacciones no covalentes, tienen libertad para moverse lateralmente en el plano de la membrana. La cara externa de la membrana plasmática, la que da al medio extracelular, presenta cadenas oligosacarídicas unidas covalentemente a lípidos o a proteínas; otras membranas celulares no presentan estos componentes glucídicos. En la Figura 11.4 se representa esquemáticamente el modelo del mosaico fluido para la estructura de la membrana plasmática.
El modelo del mosaico fluido explica la apariencia trilaminar de las membranas cuando se observan al microscopio electrónico: las dos bandas oscuras externas corresponderían a los grupos de cabeza polares (que contienen átomos más pesados y que por lo tanto retienen más los electrones), mientras que la banda clara correspondería a las colas no polares de los lípidos de membrana (formadas por átomos más ligeros y por lo tanto más transparentes a los electrones). Por otra parte, un grosor de entre 5 y 8 nm es el que cabría esperar de una bicapa de lípidos anfipáticos con proteínas incrustadas en ella y sobresaliendo a ambos lados. Los experimentos llevados a cabo con diferentes tipos de lípidos anfipáticos indican que éstos en medio acuoso tienden espontáneamente a formar estructuras como las micelas, bicapas y liposomas, ya estudiadas anteriormente, lo que también apoya la idea de una bicapa lipídica como elemento básico estructural en las membranas celulares.
El carácter fluido de las membranas se estableció mediante el marcaje de moléculas individuales de lípidos y su posterior seguimiento en el seno de la bicapa. Así pudo comprobarse que los movimientos de los lípidos dentro de cada monocapa son muy frecuentes y rápidos, mientras que son mucho más raros los cambios de una a otra monocapa (difusión "flip-flop").
El grado de fluidez de las membranas depende de la temperatura y de la composición en ácidos grasos de sus lípidos constituyentes. A temperaturas muy bajas los lípidos de membrana tienden a adoptar un ordenamiento casi cristalino (paracristalino). Por encima de una temperatura que es característica de cada membrana las moléculas de los lípidos comienzan a moverse y la membrana pasa al estado fluido. La temperatura de transición del estado paracristalino al estado fluido depende de la composición lipídica; los ácidos grasos saturados favorecen un mayor empaquetamiento de los lípidos de membrana y por lo tanto la temperatura de transición será mayor cuanto mayor sea la proporción de éstos ácidos grasos en la membrana. Por el contrario, los cambios de orientación existentes en las cadenas de los ácidos grasos insaturados dificultan el empaquetamiento de los lípidos de membrana en un ordenamiento paracristalino, por lo tanto la temperatura de transición será menor cuanto más abundante sea este tipo de ácidos grasos. Esta es la razón por la que en los organismos homeotermos, que mantienen una temperatura corporal constante y elevada, abundan los lípidos de membrana ricos en ácidos grasos saturados, mientras que en los organismos poiquilotermos, incapaces de regular su temperatura corporal, son más abundantes los lípidos de membrana ricos en ácidos grasos insaturados. Se ha comprobado que distintos tipos de células mantenidos en cultivo son capaces de alterar la composición en ácidos grasos de sus lípidos de membrana respondiendo a las variaciones de la temperatura ambiente, todo ello con el objeto de mantener la fluidez de sus membranas celulares. Por otra parte, el colesterol y esteroles afines, debido a la rigidez del sistema de anillos condensados de su molécula, tienden a impedir, actuando a modo de cuña, la agregación de los demás lípidos de membrana en ordenamientos paracristalinos, con lo que también colaboran en el mantenimiento del estado fluido (Figura 11.4).
El carácter fluido de las membranas es de una gran importancia biológica. Por una parte este carácter fluido permite que la membrana, y con ella la célula, sufra deformaciones que son la base de muchos movimientos celulares. Por otra, posibilita que tanto la membrana plasmática, como las de diferentes orgánulos puedan sufrir fenómenos de fusión y escisión del tipo de los que se representan en la Figura 11.5; estos fenómenos, en los que porciones de membrana se desprenden en forma de vesículas para luego fundirse con otras membranas, constituyen la base la base de una compleja relación dinámica entre los distintos compartimentos celulares que se conoce con el nombre de flujo de membrana y será estudiada con más detalle en otro capítulo.
En cuanto a las proteínas que forman parte de la membrana se distinguen dos tipos según su mayor o menor grado de asociación con la bicapa lipídica (ver Figura 11.6):
1) Proteínas integrales (o intrínsecas).- Se encuentran íntimamente asociadas a la bicapa por lo que resulten difíciles de extraer de la misma. Una parte sustancial de su molécula se encuentra sumergida en la bicapa lipídica estableciéndose interacciones hidrofóbicas entre los grupos R de los restos de aminoácidos no polares y las colas hidrocarbonadas de los lípidos. Se pueden extraer de la bicapa utilizando detergentes, que interfieren con dichas interacciones hidrofóbicas; cuando se realiza la extracción tienden a precipitar una vez eliminado el detergente, es decir, son poco solubles en agua. Algunas proteínas integrales atraviesan la membrana de lado a lado (proteínas transmembrana) y otras lo hacen sólo en parte.
En los últimos años se ha podido determinar la conformación tridimensional de unas cuantas proteínas integrales; todas ellas presentan dos dominios bien diferenciados: en uno de ellos, el que se encuentra sumergido en la bicapa, los restos de aminoácidos hidrofóbicos se sitúan exteriormente, de manera que puedan interactuar con las colas hidrocarbonadas de los lípidos; en el otro, formado por las zonas de la molécula que asoman a uno y otro lado de la bicapa, son los restos de aminoácidos polares los que se encuentran situados en superficie, de manera que quedan expuestos al agua. Este tipo de conformación tridimensional contrasta con el mucho más común que presentan las proteínas globulares solubles, las cuales tienen todos sus restos polares en superficie y los no polares sepultados en el núcleo proteico.ico.
2) Proteínas periféricas (o extrínsecas).- Su grado de asociación con la bicapa es mucho más débil. Se encuentran unidas a las cabezas polares de los lípidos o bien a proteínas integrales mediante interacciones débiles. Pueden liberarse de la membrana con tratamientos relativamente suaves (cambios en el pH o fuerza iónica) que rompen estas interacciones; una vez liberadas son completamente hidrosolubles. Algunas proteínas periféricas están ancladas covalentemente a lípidos de la bicapa y pueden ser liberadas por acción de enzimas específicos. Las proteínas periféricas presentan conformaciones tridimensionales que en general no difieren de las de otras proteínas globulares.
La composición y estructura supramolecular de las membranas celulares, que acabamos de estudiar, hacen que éstas, lejos de constituir meras barreras pasivas, participen de una manera activa en la regulación del tráfico de sustancias entre la célula y su entorno y entre los distintos compartimentos celulares.
Tomado de CURSO DE BIOLOGÍA Alejandro Porto Andión http://www.bionova.org.es/biocast/p2temas.htm todos los derechos reservados, se copia con fines de educación y sin fines de lucro
Actividad 4 "Virus"
El término virus era utilizado en la antigua Roma para referirse a cualquier veneno de origen animal, y las enfermedades producidas por estos venenos eran conocidas como virulentas.
Cuando a finales del siglo XIX se reconoció el papel de los microorganismos en la producción de enfermedades y a identificarse los gérmenes responsables de muchas de ellas, los microbiólogos comenzaron a utilizar el término virus para designar a todos los microorganismos patógenos.
Así, en esta época se consideraba que las bacterias eran los “virus” causantes de las enfermedades. Existían sin embargo algunos gérmenes infecciosos que se resistían a su identificación. Pasteur, aunque pudo demostrar que la rabia era producida por un agente infeccioso específico y transmisible, fue incapaz de cultivar este agente en los medios de cultivo en los que habitualmente crecían las bacterias, y tampoco pudo visualizar al microscopio ningún ejemplar al que poder atribuir la enfermedad. Poco después se pudo comprobar que estos agentes infecciosos tan escurridizos debían tener un tamaño mucho menor que el de las bacterias, ya que eran capaces de atravesar filtros de porcelana en cuyos finos poros quedaban retenidas todas las bacterias conocidas.
La investigación sobre los virus sufrió un impulso considerable con la con la entrada en escena, a finales de la década de 1930, de un grupo de investigadores procedentes de campo de las ciencias físicas, capitaneados por Max Delbrück, que habían decidido dirigir sus esfuerzos a averiguar la naturaleza del material genético. Estos investigadores encontraron en los virus bacteriófagos un material experimental extraordinariamente útil para sus propósitos. Desde entonces el desarrollo del conocimiento de la naturaleza y modo de vida de los virus corrió parejo y sirvió de apoyo al de los conocimientos en los campos de la bioquímica y la genética molecular.
NATURALEZA DE LOS VIRUS.Los virus son entidades subcelulares, es decir, su grado de organización es inferior al celular. Están constituidos casi a partes iguales por proteínas y un ácido nucleico, que puede ser DNA o RNA pero nunca los dos a la vez. Son capaces de penetrar en las células vivas y de reproducirse en su interior y sólo allí, por lo que puede considerárseles parásitos intracelulares obligados. Sin embargo, el tipo de parasitismo que desarrollan los virus presenta características que lo hacen diferente de cualquier otro conocido, pues tiene lugar a nivel genético.
Los virus carecen de maquinaria metabólica propia y en su lugar utilizan la de la célula parasitada. Cuando un virus penetra en una célula toma el control de su metabolismo de manera que una parte de los enzimas y de la maquinaria celular de producción de energía abandona sus funciones primordiales y se ponen al servicio del virus, dedicándose exclusivamente a la producción de la progenie viral. Las nuevas partículas víricas así generadas tienen a su vez capacidad infectiva y pueden penetrar en otras células para reproducirse en su interior.
Este particular modo de vida que exhiben los virus ha suscitado extensas discusiones acerca de si deben ser considerados o no como auténticos seres vivos. En efecto, en tanto que un virus se encuentra reproduciéndose en el interior de una célula viva exhibe al menos una de las funciones que tradicionalmente se vienen considerando características de los seres vivos, es decir, la reproducción. Sin embargo, cuando se encuentran fuera de las células, las partículas virales no presentan ningún tipo de actividad bioquímica, carecen de un metabolismo energético propio, e incluso pueden cristalizar a partir de suspensiones y los cristales resultantes permanecer inactivos durante largos períodos sin perder su estructura y propiedades, comportándose a todos los efectos como materia inanimada. Cuando los virus cristalizados se suspenden de nuevo en un medio adecuado y se permite su acceso a células vivas recuperan su capacidad infectiva y vuelven a reproducirse en el interior de las mismas.
Algunos autores han tratado de zanjar la polémica argumentando que los virus se encuentran “en la frontera de la vida” y que sólo deben considerarse seres vivos cuando están reproduciéndose en el interior de las células parasitadas. Es posible que se trate de una de esas discusiones bizantinas que salpican la historia de la ciencia. Lo que sí se puede afirmar es que los virus constituyen uno de los productos más sofisticados de la evolución biológica en la medida en que se encuentran entre los que con mayor eficacia y economía manipulan la materia y la energía del entorno en su propio beneficio.
ESTRUCTURA DE LOS VIRUS.
Las partículas víricas individuales con capacidad infecciosa se denominan viriones. Un virión se compone de una molécula de ácido nucleico (DNA o RNA) y una cubierta proteica que la envuelve denominada cápside. Su tamaño oscila entre los 20 y los 300 nm.

El ácido nucleico constituye el genoma del virus y contiene información para la síntesis de las proteínas de la cápside y, en algunos casos, para la de algunos enzimas implicados en la replicación del propio ácido nucleico y en la expresión de su información. Los ácidos nucleicos virales, tanto en el caso de los virus de DNA como en el de los de RNA, son en unos casos monocatenarios y e otros bicatenarios. En muchos virus de DNA la molécula presenta estructura circular, aunque también los hay de estructura lineal. Por el contrario entre los virus de RNA predominan los de estructura lineal aunque se han descrito algunos casos de estructura circular. El tipo de ácido nucleico y sus características estructurales constituyen uno de los principales criterios de clasificación de los virus.
La cápside viral está constituida por proteínasglobulares denominadas capsómeros que espontáneamente se asocian para formar una estructura tridimensional hueca que alberga al ácido nucleico en su interior. Por lo general, las cápsides virales presentan formas geométricas regulares que responden a alguno de los siguientes tipos:
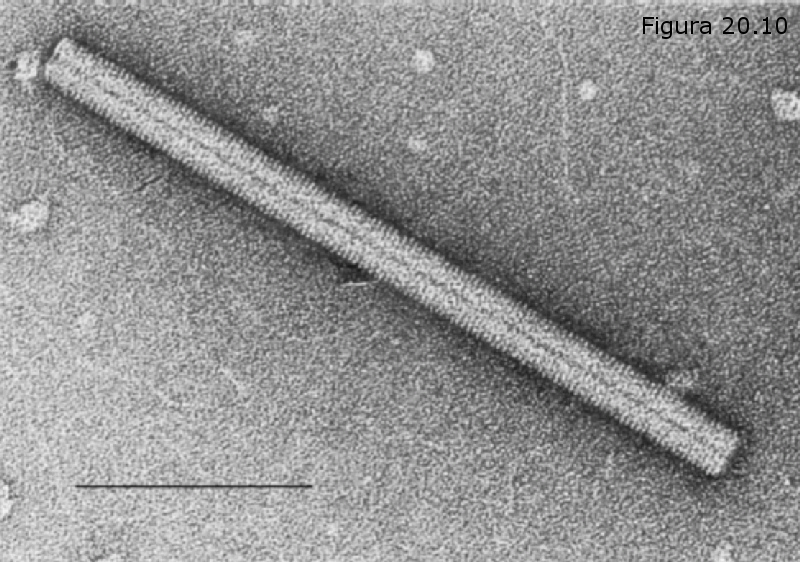
Cápside helicoidal.- Los capsómeros son todos iguales entre sí y se disponen helicoidalmente alrededor de un armazón que no es otro que el propio ácido nucleico viral. Es el caso del virus del mosaico del tabaco (Figura 20.11).
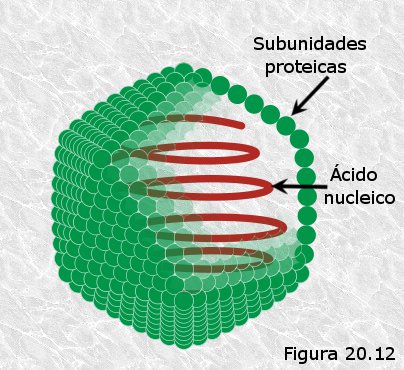
Cápside icosaédrica (Figura 20.12).- Existen al menos tres tipos de capsómeros que se disponen ocupando respectivamente las caras, aristas y vértices de un icosaedro regular hueco, en cuyo interior se empaqueta el ácido nucleico. Muchos virus presentan tipos adicionales de capsómeros que se disponen en el exterior de la estructura y tienen la misión de interactuar con la superficie de las células a infectar. Entre los virus de cápside icosaédrica se encuentran los adenovirus responsables del catarro común.

Cápside compleja.- Presentan distintos tipos de capsómeros de cuyo ensamblaje resultan las siguientes estructuras que forman parte de la cápside: a) Cabeza.- estructura icosaédrica similar a las cápsides icosaédricas ya comentadas; b) Cola.- estructura helicoidal hueca que comunica la cabeza con el exterior y permite la salida del ácido nucleico durante la infección; c) Collar.- anillo proteico que ensambla la cabeza y la cola; d) Placa basal.- estructura situada al final de la cola que permite al virus fijarse sobre la superficie de la célula a infectar; puede presentar unas prolongaciones denominadasespinas que facilitan la adsorción de la partícula viral a la superficie celular. Muchos virus bacteriófagos presentan este tipo de cápside (Figura 20.13).
Además de los componentes estructurales de la cápside algunos viriones incluyen algunas moléculas proteicas adicionales, con función enzimática, que desempeñan diferentes funciones en el proceso de infección. Tal es el caso del virus de la inmunodeficiencia humana, responsable del SIDA, cuyo virión contiene una molécula del enzima transcriptasa inversa, necesaria durante la fase inicial de la infección.
CLASIFICACIÓN DE LOS VIRUS.
El principal criterio que se ha utilizado para clasificar a los virus es el tipo de ácido nucleico que presentan. En la década de 1970 el experto virólogo y premio Nobel David Baltimore propuso un sistema de clasificación de los virus de una gran sencillez y elegancia, que, con pequeñas modificaciones, sigue utilizándose hoy en día. El sistema se basa en que la expresión del genoma viral en forma de proteínas virales siempre pasa por una molécula de mRNA. Los pasos que se han de ejecutar para sintetizar esta molécula a partir de un genoma viral concreto permite establecer seis grupos principales (Figura 20.14),en función de que el ácido nucleico sea DNA o RNA, de que éste sea de cadena doble o cadena sencilla, y, para los virus de RNA, de cual sea el modelo de expresión de la información genética que poseen (transcripción ordinaria o transcripción inversa).

En segundo lugar, para establecer los distintos grupos dentro de los seis principales, se utilizan criterios estructurales como la presencia o ausencia de envoltura lipoproteica o la morfología de la cápside viral. Por último, se recurre al tipo de células a las que infectan (animales, vegetales o bacterianas). Así se han establecido alrededor de 30 grupos de virus diferentes.
Tomado de CURSO DE BIOLOGÍA Alejandro Porto Andión http://www.bionova.org.es/biocast/p2temas.htm todos los derechos reservados, se copia con fines de educación y sin fines de lucro
Suscribirse a:
Entradas (Atom)